Aetiology
CGD is caused by mutations in genes encoding sub-units of the NADPH oxidase (nicotinamide adenine dinucleotide phosphate-oxidase) complex. X-linked CGD is caused by genetic defects of the gene encoding the gp91phox NADPH oxidase sub-unit, located on chromosome Xp21.1; it accounts for around 65% of CGD.[1][2][18] Rarely, females with skewed X-inactivation patterns can present with X-linked CGD.
The autosomal recessive form is due to mutations in genes encoding four of the NADPH oxidase sub-units: p22phox, p47phox, p67phox, or p40phox, located on chromosomes 16, 7, 1, and 22, respectively.[1][2][19] Mutations of the p47phox sub-unit gene constitute the most common autosomal recessive cause (25% of CGD), followed by defects in the p67phox and p22phox sub-units (around 5% of CGD each), and the p40phox sub-unit (rare).[2][18] Autosomal recessive forms affect both sexes.[18] Mutations in EROS (CYBC1/C17ORF62), which regulates abundance of the gp91phox-p22phox heterodimer, can also cause CGD.[20]
Pathophysiology
The NADPH oxidase complex is used by phagocytes (neutrophils and mononuclear phagocytes such as macrophages) to generate superoxide, important for effective microbial killing by phagocytes.[21] Two sub-units, gp91phox and p22phox, exist together within phagocyte plasma and granule membranes and form the flavocytochrome b558. Three other sub-units of the NADPH oxidase complex exist together in the cytosol: p40phox, p47phox, and p67phox. This cytosolic complex localises to the cytochrome b558 structure after phagocyte activation, along with Rac2. This complete complex generates superoxide, using NADPH as the electron donor. Electrons are transported across the phagosome membrane and on to oxygen within the phagosome.
Owing to the defect in NADPH activity, phagocytes are unable to generate superoxide. Superoxide, produced by the NADPH oxidase complex, is postulated to aid in initially alkalinising the phagosome pH. Coupled with compensatory ion fluxes in response to an electronegative charge generated across the vacuole membrane, the alkalinised pH optimises conditions for microbicidal proteases (e.g., elastase, cathepsin G) released by primary granules.[22][23] This theory helps to explain the susceptibility to infection with catalase-negative organisms such as (artificially modified) Staphylococcus aureus and Aspergillus nidulans in mouse models.[24][25] Failure to undergo oxidative burst also renders neutrophils unable to form neutrophil extracellular traps, which further impairs their antimicrobial activity.[26] Repeated inflammation can cause extensive granuloma formation in the skin, lungs, lymph nodes, liver, or bones, which can obstruct the gastrointestinal or genitourinary tracts. Patients with residual NADPH oxidase activity have an improved clinical course, irrespective of the subtype.[27][Figure caption and citation for the preceding image starts]: NADPH oxidase complexCreated by previous contributor, Rebecca A. Marsh, MD; used with permission from Cincinnati Children’s Hospital Medical Center Diagnostic Immunology Laboratory [Citation ends].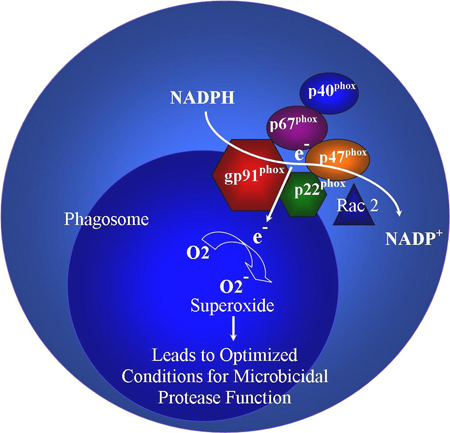
Use of this content is subject to our disclaimer