Article Text

Abstract
Background: The purpose of this study was to use laser scanning in vivo confocal microscopy to elucidate the location and morphology of stromal nerves in the normal human central cornea.
Methods: Analysis was performed via an established database of laser-scanning in vivo confocal microscopy on images of the central cornea of normal subjects. The depth and morphology of the stromal nerves were determined.
Results: The population of this study consisted of 99 eyes of 99 healthy subjects (38 male, 61 female). The mean age of the group was 34.7 (SD 13.3, range 13–84) years. Two morphologically different populations of stromal nerves were observed: (1) straight, dichotomous branching nerves; and (2) tortuous nerves with a beaded appearance. The mean recorded depth of straight stromal nerves (186 (SD 66) μm) was significantly deeper than the mean depth of the tortuous stromal nerves (140 (SD 87) μm) (p<0.001).
Conclusions: The current study identified two morphologically distinct stromal nerve populations in the normal human cornea. We hypothesise that the two morphological nerve populations described here may represent functionally heterogeneous nerves. Further research is required to determine if these in fact represent different types of sensory nerves.
Statistics from Altmetric.com
In vivo confocal microscopy is a non-invasive method used to analyse the human living cornea. This technique has led to a greater understanding of the architecture of corneal nerves.1–3
A number of reports have described the presence of tortuous, “aberrant” nerve trunks in the anterior and mid corneal stroma in diseases such as Schnyder’s crystalline dystrophy,4 type 2 diabetes mellitus,5 leprosy,6 and amiodarone-induced keratopathy,7 and following photorefractive keratectomy.8
Only a few studies have examined stromal nerves in the normal “healthy” population. Indeed, most of these studies have been performed on post mortem tissue9–11 and only one published study has used in vivo confocal microscopy to analyse stromal nerve architecture in the normal cornea.2 The recent development of laser-scanning in vivo confocal microscopy (LSCM) has led to improved imaging of the epithelium, sub-basal nerve plexus and stroma, producing images with greater contrast than those generated by white light in vivo confocal microscopes.12
The purpose of this study was to use LSCM to elucidate the location and morphology of stromal nerves in the normal human central cornea.
MATERIAL AND METHODS
The analysis was performed using a large database of LSCM images obtained from normal human subjects assessed between March 2005 and September 2007. LSCM had been performed on the central cornea of all subjects using the Heidelberg Retina Tomograph II Rostock Corneal Module (RCM; Heidelberg Engineering, Heidelberg, Germany) at the Department of Ophthalmology, University of Auckland, as described previously.14
The exclusion criteria were previous contact lens wear, ocular surgery or ocular trauma, the presence of other ocular diseases, and the presence of systemic diseases that might affect the cornea. Images from one cornea of each patient were selected for analysis and the eye to be examined was selected randomly.
For each subject, demographic data were recorded and all images containing stromal nerves were included. The location depth (μm), as established by the in vivo confocal microscope, was recorded for each image that contained stromal nerves. All images selected were subsequently randomised by a single observer (DVP) and then a qualitative description of the nerves was established by an independent observer (NV).
RESULTS
The population of this study consisted of 99 eyes of 99 healthy subjects (38 male, 61 female). The mean age of the group was 34.7 (SD 13.3, range 13–84) years. There were no visible stromal nerves in 14 corneas from 14 subjects. From the remaining subjects, 462 images exhibited stromal nerves and were selected for further analysis.
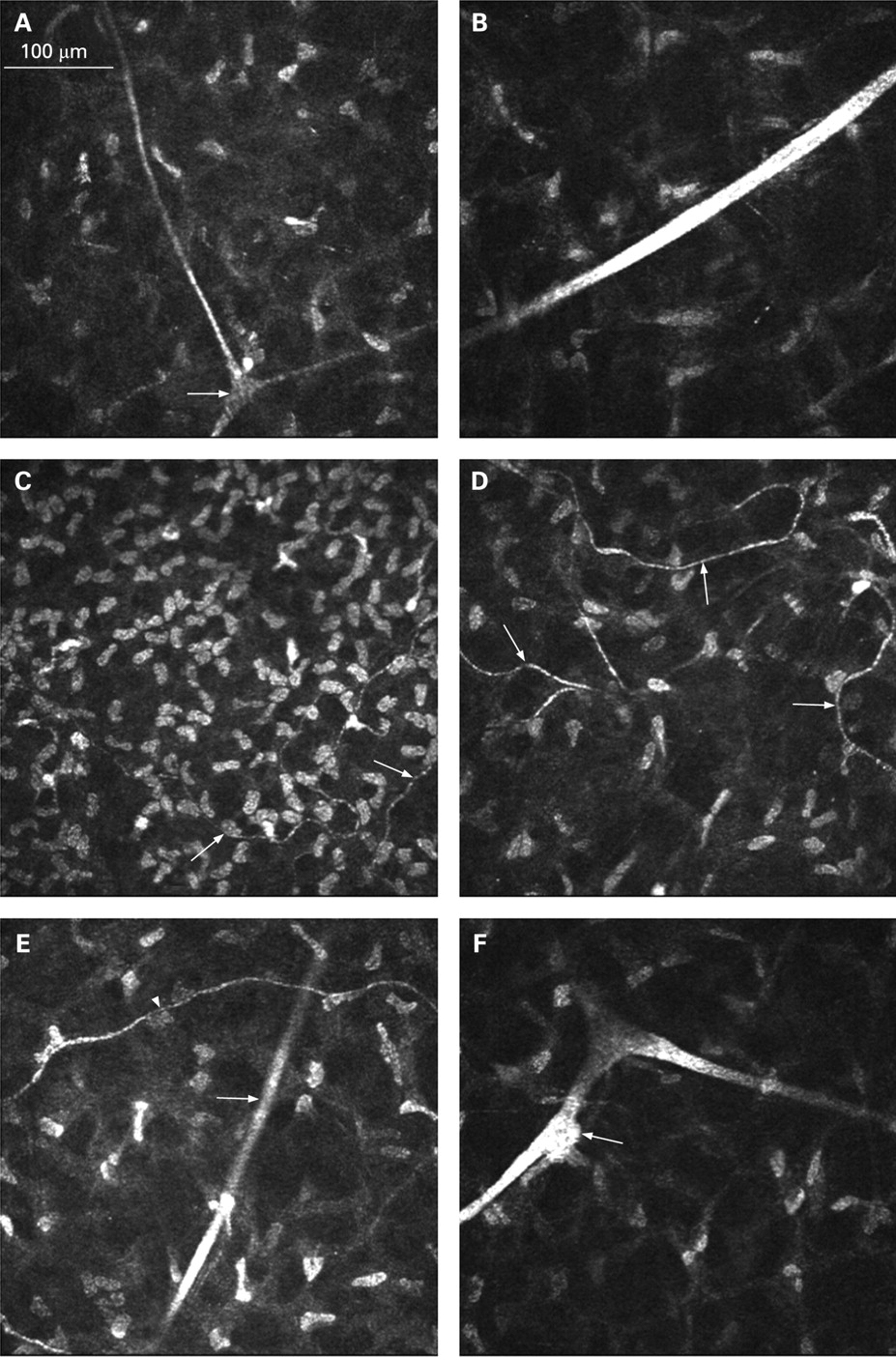
Within the corneal stroma two distinct nerve populations could be distinguished: (1) straight nerves, often with dichotomous branching (fig. 1A, B); and (2) tortuous nerves with a beaded appearance (fig. 1C, D).

{kind=link}
Laser-scanning in vivo confocal microscopy images of corneal stromal nerves in normal subjects. (A) Straight dichotomous branching (arrow) nerve in the anterior part of the stroma (depth 130 μm). (B) Thick straight nerve in the mid stroma (depth 350 μm). (C) Tortuous nerves (arrows) in the anterior part of the stroma (depth 70 μm). (D) Tortuous nerves (arrows) in the mid stroma (depth 210 μm). (E) Straight nerve (arrow) and tortuous nerve (arrowhead) (depth 160 μm). (F) Keratocyte (arrow) apparently in contact with a stromal nerve (depth 290 μm).
Straight nerves were present in 80 subjects, while tortuous nerves were present in 43 subjects.
Tortuous nerves were most often observed in the anterior stroma and mid stroma. Occasionally, these nerves were noted alongside straight nerves (fig. 1E).
Interestingly, some stromal nerves were occasionally in apparent contact with keratocytes (fig. 1F). No nerves were visible in the posterior corneal stroma.
The mean recorded depth of straight stromal nerves (186 (SD 66) μm) was significantly deeper than the mean depth of the tortuous stromal nerves (140 (SD 87) μm) (p<0.001, Mann–Whitney U test).
DISCUSSION
The current study identified two morphologically different corneal stromal nerve populations. The observation of thick, straight, stromal nerves, often with a dichotomous branching pattern, is in accordance with data from previous research.2 14 15 However, a second morphologically distinct nerve population, consisting of thinner, tortuous nerves with a beaded appearance, was observed in the corneal stroma and these have not been described in the literature before.
In the classic anatomical description of human corneal nerve architecture, thick nerve bundles originating from the ophthalmic division of the trigeminal nerve enter the peripheral stroma in a radial fashion and run anteriorly towards the central cornea. Di- or trichotomous branching occurs to give rise to smaller branches that innervate the anterior and mid stroma.2 The sub-epithelial plexus is formed immediately beneath Bowman’s layer and has been described as being limited to the peripheral cornea and possibly absent from the central cornea.14 Oliveira-Soto and Efron were able to image the sub-epithelial nerve plexus in the central cornea using slit-scanning in vivo confocal microscopy (SSCM): they reported that these nerves showed great variability in tortuosity and noted that 50% of these nerves exhibited beading.2 Although this suggests that the tortuous nerve population described in the current study may represent the sub-epithelial plexus, this plexus has been described as only being present in a single plane in the anterior stroma immediately posterior to Bowman’s layer,2 16 whereas tortuous beaded nerves were detected in locations as deep as the mid stroma in the current study.
Although there are no published data validating corneal depth measurements using the RCM, the errors produced by the combination of involuntary anterior–posterior movement of the subject and the requirement of manual adjustment of the focal plane mean that this method is unlikely to have the degree of accuracy and repeatability of confocal microscopy through focusing (CMTF)17 or the ConfoScan with the Z-ring adapter.18 However, despite this significant limitation, the depth measurements in this study were generally in accordance with the corneal depth associated with preceding, and subsequent scans and subjective examination of the acquired images clearly confirms the presence of tortuous beaded nerves in the anterior and mid-stromal regions (fig. 1).
There were no visible stromal nerve images in 14% of central corneas in this study. Because all subjects in the study were considered to have normal corneas on history and examination, it can be presumed that stromal nerves were present in all corneas but had not been detected in some. This is probably due to a combination of the relatively low density of stromal nerves in the central cornea2 and the inconsistency in the total number of images acquired for each subject in this study.
One hypothesis regarding the two morphological nerve populations described here is that they may represent functionally heterogeneous nerves. It is known that the human cornea contains different functional types of corneal sensory fibres: mechano-nociceptors that fire in response to indentation of the corneal surface, polymodal-nociceptors that are activated by mechanical stimuli, heat and chemical irritants, and cold-sensitive receptors.19 Elegant three-dimensional reconstructions by Guthoff et al16 have shown that Aδ and C fibres penetrate Bowman’s layer orthogonally and separately and both types of nerve fibres are reported to arise from the sub-epithelial nerve plexus. An alternative theory is that the observations described in the current study are the result of a single population of nerve fibre bundles that become morphologically different to facilitate nerve passage through the more anterior stromal layers. A possible reason for more beading near the anterior cornea is that the extended tortuous length of the nerves in the anterior stroma requires periodic “power stations” (dense accumulations of mitochondria and other organelles) to facilitate efficient nerve conduction.
In the current study, keratocytes were occasionally noted to be in apparent contact with stromal nerves (straight or tortuous) or vice versa. Interestingly, in a detailed electron microscopic analysis of nerve fibres in the human cornea, keratocytes have been shown to occasionally invaginate nerve fibres.15 Contact between keratocytes and nerves is also observed more often in patients with keratoconus than in control subjects,20 and it has been suggested that corneal nerve–keratocyte interactions in keratoconus may lead to remodelling of the extracellular matrix.21
The majority of previous in vivo confocal microscopy studies describing nerve morphologies used either a tandem-scanning confocal microscope or an SSCM, while in this study an LSCM was used. The LSCM has been shown to generate higher quality images of the sub-basal nerve plexus and the corneal stroma than the SSCM. The images produced by a LSCM have significantly greater contrast than those produced by SSCM; the contrast in LSCM images remains high up to all edges of each image while the contrast decreases remarkably towards the lateral edges of each SSCM image.12 The improved resolution and contrast of the LSCM in this study may thus have enabled the visualisation of this second population of thinner, tortuous, beaded nerves in the corneal stroma.
Stromal nerve diameter and density were not measured in this study. Stromal nerve diameter measurements would be inaccurate since these nerves commonly traverse obliquely, relative to the en face section of the images. The cross-section is therefore not always through the centre of the nerve, and so off-centre cross-sections will make the nerve appear falsely thinner. Stromal nerve orientation is also an important factor when considering nerve density analysis, unlike analysis of sub-basal plexus nerve density.1 Visible nerve length per frame area will vary with the path of the nerve through the field, but it does not necessarily relate to nerve density in the stroma.
In conclusion, the current study identified two apparently morphologically distinct stromal nerve populations in the normal adult human cornea. Further research is obviously required to determine if these in fact represent different types of sensory nerves. Furthermore, a prospective study including corneal aesthesiometry and measurements of the central corneal thickness (to enable localisation of the stromal nerves to anterior, mid or posterior stromal locations) would be useful in providing further insight into the effects of corneal disease on morphology and function of corneal nerves.
REFERENCES
Footnotes
Competing interests: None declared.
Ethics approval: Obtained.
Patient consent: Obtained.