Article Text

Abstract
Objective Hepatitis B virus-related acute-on-chronic liver failure (HBV-ACLF) pathophysiology remains unclear. This study aims to characterise the molecular basis of HBV-ACLF using transcriptomics.
Methods Four hundred subjects with HBV-ACLF, acute-on-chronic hepatic dysfunction (ACHD), liver cirrhosis (LC) or chronic hepatitis B (CHB) and normal controls (NC) from a prospective multicentre cohort were studied, and 65 subjects (ACLF, 20; ACHD, 10; LC, 10; CHB, 10; NC, 15) among them underwent mRNA sequencing using peripheral blood mononuclear cells (PBMCs).
Results The functional synergy analysis focusing on seven bioprocesses related to the PBMC response and the top 500 differentially expressed genes (DEGs) showed that viral processes were associated with all disease stages. Immune dysregulation, as the most prominent change and disorder triggered by HBV exacerbation, drove CHB or LC to ACHD and ACLF. Metabolic disruption was significant in ACHD and severe in ACLF. The analysis of 62 overlapping DEGs further linked the HBV-based immune-metabolism disorder to ACLF progression. The signatures of interferon-related, neutrophil-related and monocyte-related pathways related to the innate immune response were significantly upregulated. Signatures linked to the adaptive immune response were downregulated. Disruptions of lipid and fatty acid metabolism were observed during ACLF development. External validation of four DEGs underlying the aforementioned molecular mechanism in patients and experimental rats confirmed their specificity and potential as biomarkers for HBV-ACLF pathogenesis.
Conclusions This study highlights immune-metabolism disorder triggered by HBV exacerbation as a potential mechanism of HBV-ACLF and may indicate a novel diagnostic and treatment target to reduce HBV-ACLF-related mortality.
- liver failure
- hepatitis B
- cirrhosis
Data availability statement
Data, that is raw reads from RNA-seq experiments in fastq files, are available in a public, open access repository without restrictions on the use or distribution of the data. The Sequence Read Archive database project accession number for accessing the data produced for this study is PRJNA548207 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA548207/).
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Acute-on-chronic liver failure (ACLF) has regional phenotypic specificities due to the disease aetiology and precipitating event.
Hepatitis B virus-related ACLF (HBV-ACLF) exhibits clinical characteristics different from alcoholic liver disease-related ACLF in Western populations.
The transcriptomic-based molecular mechanism underlying disease development and the prognosis of HBV-ACLF remains unclear.
What are the new findings?
The transcriptome of HBV-ACLF is characterised by an excessive immune response triggered by HBV exacerbation that drives chronic hepatitis B or liver cirrhosis to acute-on-chronic hepatic dysfunction and ACLF.
Dysregulation of immune responses leads to metabolic disorder and inflammation and induces multiorgan failure in patients with HBV-ACLF.
Immune-metabolism disorder is a core axis of disease development and progression in patients with HBV-ACLF.
Four differentially expressed genes (PPAR gamma, MER tyrosine kinase, thrombospondin 1 and semaphorin 6B) are identified as potential biomarkers that reflect dysregulated pathways underlying the virus-based immune-metabolism disorder in patients with ACLF.
How might it impact on clinical practice in the foreseeable future?
The new findings may direct the development of effective early diagnostic and treatment strategies targeting immune regulation and metabolic balance to reduce the high mortality rate of patients with HBV-ACLF.
Introduction
Acute-on-chronic liver failure (ACLF) develops in patients with chronic liver diseases, regardless of the presence of cirrhosis and represents the common final pathway leading to death.1 An understanding of the molecular mechanism of ACLF pathogenesis is essential for the development of effective early diagnostic and treatment strategies. The chronic liver failure (CLIF) Consortium Acute-On-Chronic Liver Failure in Cirrhosis (CANONIC) study first developed the definition and diagnostic criteria for ACLF in patients with acutely decompensated alcoholic liver disease or hepatitis C virus-related cirrhosis.2 This well-accepted definition recognises ACLF as a distinct syndrome based on the presence of organ failure and a high mortality rate. Our recent large prospective multicentre study (Chinese Group on the Study of Severe Hepatitis B, COSSH) documented that hepatitis B virus-related ACLF (HBV-ACLF) exhibits clinical characteristics different from alcoholic liver disease-related ACLF in Western populations, and we proposed the COSSH-ACLF criteria for the diagnosis of ACLF in the HBV-positive population, as these criteria exhibited improved diagnostic sensitivity and prognostic accuracy.3
Hepatocyte injury and death are known to initiate ACLF; however, the cause of hepatocyte injury and the pathway involved in ACLF progression remain unclear.4 The CANONIC study indicated that an intense systemic inflammatory response is the main cause of acute deterioration in patients with alcoholic liver disease-related and hepatitis C virus-related ACLF.5 6 As mentioned earlier, the two largest prospective multicentre studies of ACLF to date, CANONIC and COSSH, revealed regional phenotypic specific differences in the disease aetiology and precipitating events of ACLF.7 Nevertheless, immune cells appear to play a crucial role in ACLF; however, the ongoing processes in these cells may be different and have not been investigated in depth to date. In the present study, we aim to provide a transcriptomic-based dataset from circulating immune cells (peripheral blood mononuclear cells, PBMCs) collected at the different stages of disease progression in patients with HBV-ACLF to clarify the detailed molecular mechanisms underlying disease development and their effects on the liver tissue and prognosis of patients with HBV-ACLF, as well as to develop and validate potential biomarkers to improve the diagnosis and prognosis of HBV-ACLF.
Materials and methods
Study design
To elucidate the detailed transcriptomic-based molecular mechanism of the development and progression of HBV-ACLF, we prospectively and randomly selected 340 patients with HBV aetiology from the COSSH study open cohort to evaluate and characterise the evolving disease course starting from chronic hepatitis B (CHB, n=70) or liver cirrhosis (LC, n=70) and progressing through acute-on-chronic hepatic dysfunction (ACHD, n=70) to ACLF (n=130). Relevant clinical and follow-up data were collected from the electronic data capture system and case report forms. Normal, healthy volunteers were enrolled as normal controls (NC, n=60). Antiviral nucleoside analogues were immediately administered if HBV DNA was detected in patients with ACLF or ACHD according to the Consensus Recommendations of the Asian Pacific Association for the Study of the Liver (2009).8 The patients and controls were randomly allocated into a sequencing group and a validation group (figure 1A). Among the sequencing group, 20 patients with HBV-ACLF from the COSSH cohort were selected using stratified random sampling according to the prevalence of ACLF grades in the COSSH study (ACLF-1, 60.6%; ACLF-2, 33.0% and ACLF-3, 6.4%).3 PBMCs from 65 subjects (ACLF, 20; ACHD, 10; LC and CHB, 10 each, respectively and NC, 15) in the sequencing group were subjected to transcriptomic sequencing. A functional synergy analysis (figure 1B) was conducted to identify variations in genes and biological processes related to the clinical pathophysiology of HBV-ACLF.9 Potential biomarkers were validated by quantitative reverse transcription-PCR (qRT-PCR) in PBMCs from remaining 335 subjects and immunohistochemistry (IHC) in liver tissues from patients with HBV-ACLF and in liver tissues from a rat ACLF model, respectively.

Transcriptomic characteristics of patients with HBV-ACLF. (A) Study design and patient group allocation based on the COSSH-ACLF diagnostic criteria. (B) Flow chart of the functional synergy analysis strategy. (C) Principal component analysis of subjects in the ACLF, ACHD, LC, CHB and NC groups. (D) Number of DEGs analysed in pairwise comparisons among the five groups. (E) Venn diagram of the DEGs analysed in pairwise comparisons among subjects in the CHB, LC and NC groups. The number of mRNA sequencing (n=20/10/10/10/15, ACLF/ACHD/LC/CHB/NC groups, respectively). ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; CHB, chronic hepatitis B; COSSH, Chinese Group on the Study of Severe Hepatitis B; DEGs, differentially expressed genes; HBV, hepatitis B virus; LC, liver cirrhosis; NC, normal controls; PBMC, peripheral blood mononuclear cell.
Definitions
COSSH-ACLF and ACHD
HBV-ACLF was diagnosed based on the COSSH-ACLF criteria.3 The definition identified HBV-ACLF as a complicated syndrome with a high short-term mortality rate that develops in patients with HBV-related chronic liver disease, regardless of the presence of cirrhosis, and it is characterised by acute deterioration of liver function and hepatic and/or extrahepatic organ failure. HBV-ACLF comprises three grades: ACLF-1, ACLF-2 and ACLF-3. ACLF-1 comprises four types of patients: (1) patients presenting with liver failure alone with an international normalised ratio (INR) ≥1.5 and/or kidney dysfunction and/or hepatic encephalopathy (HE) grade I or II; (2) patients with kidney failure alone; (3) patients with failure of a single organ failure in the coagulation, circulatory or respiratory system and/or kidney dysfunction and/or HE grade I or II and (4) patients with cerebral failure plus kidney dysfunction. ACLF-2 comprises patients with failures in two organ systems, while ACLF-3 comprises patients with failures in three or more organ systems. ACHD was defined as patients with pre-ACLF who did not fulfil the criteria for the ACLF diagnosis. ACHD comprised of two groups of patients: (1) patients with severe liver injury (total bilirubin, TB ≥5 mg/dL) with previously diagnosed CHB and (2) patients with previously diagnosed cirrhosis presenting with one of five of the following precipitating events: ascites, upper gastrointestinal bleeding, HE, bacterial infection and a high level of jaundice (TB ≥5 mg/dL).
LC, CHB and NC
In this study, LC was defined as patients with stable compensated cirrhosis. Cirrhosis was diagnosed based on previous liver biopsy results, clinical evidence of previous decompensation and laboratory tests, endoscopy (oesophageal and gastric varices) and radiological imaging of portal hypertension and/or liver nodularity. Patients with a history of decompensation (ascites/HE/upper gastrointestinal haemorrhage/bacterial infection) were excluded. Detailed information about the criteria used for the cirrhosis diagnosis is provided in online supplemental materials and methods. The enrolment criteria for the patients with CHB corresponded to the 2009 American Association for the Study of Liver Diseases (AASLD) guidelines.10 Normal healthy volunteers (18–80 years) are participants with a normal physical examination. The subjects with underlying common diseases, such as diabetes mellitus and hypertension, were excluded.
Supplemental material
Methods for mRNA sequencing, qRT-PCR and histological staining
The mRNA sequencing was performed using TruSeq RNA LT Sample Prep Kit v2 (Illumina, San Diego, California, USA). The materials, reagents, primers and antibodies, and protocols for qRT-PCR, H&E/Masson’s trichrome staining and IHC used in this study are provided in online supplemental methods, online supplemental table 1 (primers) and online supplemental table 2 (antibodies).
Functional synergy analysis
Significant differentially expressed genes (DEGs) were identified and clustered into biological processes by performing a network-based functional synergy analysis (gene set linkage analysis, GSLA).11 Biological processes were defined using publicly available gene sets (Gene Ontology, GO; Kyoto Encyclopedia of Genes and Genomes, KEGG; Reactome, etc.). Interactions between related biological processes were analysed to elucidate the molecular mechanism of HBV-ACLF. A detailed description of the identification of DEGs, principal component analysis, hierarchical clustering of gene expression, sliding window analysis of DEG functions and gene expression pathway analysis is provided in online supplemental materials and methods.
Immune profile analysis
Blood transcriptional modules (BTMs) were used to further define the differences in immune profiles among patients with different disease stages. BTMs are molecular signatures constructed under different types of immunological stimuli from PBMC transcriptomics developed by Li et al,12 which were used to reduce the transcriptomic data dimensionality. Each BTM was used as a gene set for a frequency analysis, gene set enrichment analysis (GSEA),13 paired t-test and the mean value analysis. Above analyses were used to measure BTM activity.
Statistical analysis
The results of the measurements are presented as the means±SD or medians (p25, p75), unless indicated otherwise. Student’s t-test was used to compare two continuous variables if each compared group passed the Shapiro-Wilk test (p>0.05). Paired t-tests were only used to compare the member genes of BTMs between patients with different disease stages and NC. The Mann-Whitney U test was used in other scenarios to compare variables. The χ2 test was used to compare categorical variables. Multiple comparisons corrections were performed using the Benjamini-Hochberg procedure to control for type I error. Significance was defined as p<0.05.
Results
Patients and clinical characteristics
The clinical characteristics of all 400 subjects are provided in table 1. The HBV DNA level was significantly higher in the ACLF group than in the LC and CHB groups (p<0.001). The levels of laboratory indicators, including TB and INR, were significantly higher in the ACLF group than in the other four groups. Liver failure and coagulation failure were the most frequent types of organ failure occurring in the patients with ACLF, consistent with the observations in our previous study.3 The short-term (28/90 days) mortality rate of the patients with ACLF was 33.0/46.0%, which was higher than the patients with ACHD (2.9/2.9%). Among the 65 subjects in the sequencing group (figure 1A, online supplemental table 3), one sample from a patient with LC and one NC sample did not reach the minimum sequencing reads (>20 M) quality check criterion and were excluded from this study. Sequencing reads are available in the Sequence Read Archive database (accession number: PRJNA548207, online supplemental table 4).
Clinical characteristics of patients
Transcriptomic characteristics of PBMCs from patients with HBV-ACLF
The principal component and kernel density estimation analyses of gene expression in PBMCs revealed that patients with ACLF clustered together and were clearly separated from the other patients (figure 1C and online supplemental figure 1). The NC/CHB/LC group, ACHD group and ACLF group were progressively distributed along one direction (figure 1C, red line).
The pairwise differential expression analysis illustrated that the ACLF and ACHD groups had a small number of DEGs (13), and thus they were significantly different from the other groups (>297 DEGs) (figure 1D and online supplemental tables 5–14). Although ACHD did not manifest the clinical features of ACLF, it was transcriptionally considered a precursor stage of ACLF. We also performed a multigroup comparison and identified 1078 DEGs with distinct differences in expression levels in at least one group. The hierarchical clustering analysis based on the multigroup DEGs produced the same result (online supplemental figure 2). An analysis of the LC, CHB and NC groups indicated that their pairwise DEGs did not overlap, suggesting that LC and CHB were dissimilar disease bases for ACLF (figure 1E). Therefore, these two groups were separately used as controls in further analyses.
Acute exacerbation of HBV and immune-metabolism disorder in patients with ACLF
The top 500 DEGs, which were identified by four pairwise comparisons between the ACLF/ACHD/LC/CHB group and the NC group, were subjected to a functional synergy analysis of their connections to biological processes to identify significant pathophysiological variations that underlie ACLF progression. The resulting 1014 biological process terms were summarised into seven general biological processes related to the PBMC response (viral, immune, wounding, metabolic, coagulation, inflammatory and apoptotic processes). The allocation map of these biological processes is provided in online supplemental figure 3 and online supplemental table 15.
As shown in the sliding window DEG function plot (figure 2A), virus processes were associated with all disease stages (CHB, 3/17; LC, 4/35; ACHD, 5/39 and ACLF, 5/35 GO terms/DEGs). Immune processes (27/115 GO terms/DEGs) exhibited the most significant disorder in patients with ACHD, which represented the most prominent change among all seven functional processes and across all disease stages. Inflammatory processes were also associated with all stages (CHB, 3/57; LC, 1/23; ACHD, 20/70 and ACLF, 7/35 GO terms/DEGs). However, the levels of inflammatory process dysregulation were similar between patients with CHB and ACHD and were slightly improved in patients with ACLF. Apoptotic processes (7/35 GO terms/DEGs) exhibited more obvious dysregulation in patients with CHB than in patients with ACHD (1/7 GO terms/DEGs) and ACLF (4/17 GO terms/DEGs). The dysregulation of metabolic processes was observed at all disease stages and progressively increased from CHB to ACLF (CHB, 4/38; LC, 8/41; ACHD, 22/59 and ACLF, 17/69 GO terms/DEGs). Coagulation and wounding processes are closely related to coagulation failure and liver damage, respectively, which represent the known clinical manifestations of ACLF. These three processes were dysregulated in patients with ACHD and ACLF, suggesting consistency between our transcriptomic observations and known pathophysiological variations. Based on these observations, viral, immune and metabolic processes participated in the development and progression of HBV-ACLF.

Hepatitis B virus exacerbation and immune-metabolism disorder during ACLF aggravation. (A) Functional synergy analysis of the functional implications of the top 500 DEGs in pairwise comparisons between the ACLF/ACHD/LC/CHB and NC groups. (B) Network of the biological processes identified in the functional synergy analysis of 100 DEGs between the ACHD and NC groups. (C) Network of the biological processes identified in the functional synergy analysis of 100 DEGs between the ACLF and NC groups. (D) Network of the biological processes identified based on overlapping DEGs. Top panel, number of DEGs analysed in pairwise comparisons among the ACLF, ACHD, LC and CHB groups. Bottom panel, network of the biological processes identified in a functional synergy analysis of 62 overlapping DEGs. The number of mRNA sequencing (n=20/10/10/10/15, ACLF/ACHD/LC/CHB/NC groups, respectively). ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; CHB, chronic hepatitis B; DEGs, differentially expressed genes; IL, interleukin; LC, liver cirrhosis; NC, normal controls; NK, natural killer.
The top 100 DEGs identified by comparing the ACLF/ACHD group and the NC group were subjected to a GSLA to obtain insights into the most prominent biological processes that changed from ACHD to ACLF. As shown in figure 2B,C, among the most prominent biological processes that were changed in patients with ACHD and ACLF, viral, immune and metabolic processes played central roles in the biological process network. The biological processes related to viral activation (eg, the regulation of viral replication and transcription) were altered during the progression from ACHD to ACLF and associated with the increased HBV DNA level or HBV exacerbation in patients with ACLF (table 1). Immune processes (eg, innate immune activating cell surface receptor, innate immune activating signal transduction and activation of innate immune response) were significantly regulated in patients with ACHD and ACLF. Inflammatory processes activated in response to proinflammatory factors (such as interleukin (IL)-6, IL-1 and IL-2) were dysregulated in patients with ACHD, whereas this dysregulation was mostly ameliorated in patients with ACLF (only IL-8 and antigenic stimulus were observed). Dysregulation in glycogen, cholesterol efflux and high-density lipoprotein metabolism were notably observed during the transition from ACHD to ACLF. Sixty-two DEGs were consistently identified when the ACLF and ACHD groups were compared with the LC and CHB groups (figure 2 and online supplemental table 16). These DEGs were associated with the core mechanisms underlying ACLF development. The GSLA identified 27 GO terms, 14 of which were related to viral, immune and metabolic processes, indicating that the development of ACLF is strongly correlated with virus-based immune-metabolism disorder.
Innate immune response is upregulated while the adaptive immune response is downregulated across ACLF development
The analysis of immune-related BTMs revealed the substantial upregulation of genes associated with the innate immune response, whereas the expression of genes involved in the adaptive immune response was downregulated. The DEGs associated with the innate immune response were most noticeable in five modules (interferon, monocytes, neutrophils, inflammation and dendritic cells) (figure 3A). In the interferon module, the majority of the genes in the gene cluster, including the antiviral interferon signature, innate antiviral response and type I interferon response, were significantly upregulated in the CHB and ACHD groups compared with the NC group. The results of the GSEA showed that the enrichment scores of the aforementioned gene clusters were higher in the CHB and ACHD groups. The mean expression of the modules revealed the highest expression of these DEGs in the ACHD group, with significant downregulation detected in the ACLF group, indicating that the interferon modules might be an initial factor contributing to the occurrence of ACLF.

Spectra of immune responses defined by BTMs across ACLF development. (A) Innate immune response-related BTMs were activated in patients with ACLF. (B) Adaptive immune response-related BTMs were exhausted in patients with ACLF. The first column shows the frequency analysis of significant DEGs in each module. Coloured spots represent the percentage of significantly upregulated (red) or downregulated (blue) genes. The second column shows the results of the GSEA using BTMs as gene sets. The heatmap represents the Normalised Enrichment Score (NES) calculated by the GSEA for these modules. *p<0.05, **p<0.01, ***p<0.001. The third column shows the results of the paired t-test comparing member genes between patients with different disease stages and NC. The transcriptomics data were summarised into module level scores by calculating the mean values of member genes. The colour intensity and size of nodes represents the degree of perturbation. In the fourth column, line charts represent the mean expression of each module across disease stages. The number of mRNA sequencing (n=20/10/10/10/15, ACLF/ACHD/LC/CHB/NC groups, respectively). ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; BTM, blood transcriptional module; CHB, chronic hepatitis B; DEGs, differentially expressed genes; GSEA, gene set enrichment analysis; HBV, hepatitis B virus; LC, liver cirrhosis; NC, normal controls; NK, natural killer.
The genes in the monocyte modules enriched in monocytes (I/II/III/IV/surface), activated dendritic cells/monocytes, monocyte surface signature and myeloid cells/monocytes were significantly upregulated in the ACHD and ACLF groups compared with the NC group. The GSEA showed a strong clustering of the DEGs in the ACHD and ACLF groups. The paired t-test of member genes showed the most significant changes in the six BTMs, and mean expression plots of the modules showed upregulation in the ACHD and ACLF groups. Based on these results, activation of monocyte function plays a vital role in the immune response in patients with ACLF. Similarly, the BTMs associated with neutrophils, inflammation, dendritic cells and antigen presentation showed consistent changes in the ACHD and ACLF groups. Taken together, the activation of the innate immune system might be the key mechanism underlying ACLF progression.
The BTMs associated with the adaptive immune response showed contrasting changes, and the majority of the DEGs in the gene clusters were downregulated (figure 3B). The GSEA, paired t-test and mean expression plots also showed that the BTMs associated with T cells, B cells and NK cells were significantly suppressed in the ACLF group. Thus, adaptive immunity might be exhausted in patients with ACLF.
Transcriptome reveals metabolic alterations in patients with ACLF
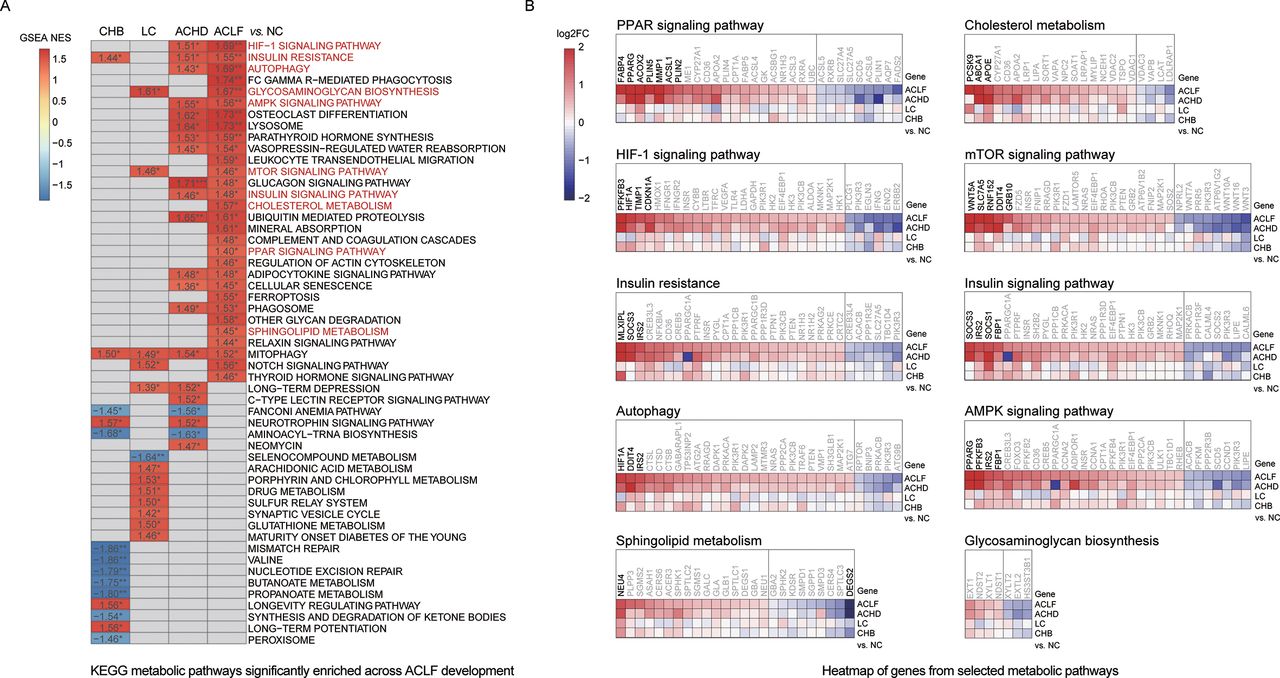
Metabolic pathways from the KEGG database were used for functional annotation based on the GSEA results to further investigate metabolic changes across ACLF development. The pathway enrichment analysis identified 53 pathways (p<0.05) that were enriched across ACLF development (figure 4A). Thirty of these pathways were upregulated in the ACLF group, including the peroxisome proliferator-activated receptor (PPAR) signalling pathway, cholesterol metabolism, hypoxia-inducible factor 1 signalling pathway, mechanistic target of rapamycin (mTOR) signalling pathway, insulin resistance, insulin signalling pathway, autophagy, AMP-activated protein kinase signalling pathway, sphingolipid metabolism and glycosaminoglycan biosynthesis. These pathways play roles in lipid metabolism, fatty acid metabolism, oxygen homoeostasis and autophagy.

Regulation of metabolism across ACLF development. (A) Metabolic pathways that were significantly altered across disease stages based on enriched KEGG pathways with only p value <0.05 are depicted here. *p<0.05, **p<0.01, ***p<0.001. Lipid handing-related and usage-related pathways are highlighted in red. Pathways in which gene expression was increased or decreased are shown in red and blue in the heatmap, respectively. The colour intensity is proportional to the NES calculated using the GSEA. (B) Heatmap of genes from selected KEGG-derived metabolic pathways. For each pathway, the top 30 DEGs are shown (adjusted p value <0.05). (All DEGs are shown if the number of detected genes is <30.) Genes with higher (red) or lower (blue) expression are proportionally displayed. Significantly differentially expressed genes (|log2-fold change|>1.5) are shown in bold. The number of mRNA sequencing (n=20/10/10/10/15, ACLF/ACHD/LC/CHB/NC groups, respectively). ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; AMPK, AMP-activated protein kinase; CHB, chronic hepatitis B; DEGs, differentially expressed genes; GSEA, gene set enrichment analysis; HBV, hepatitis B virus; LC, liver cirrhosis; NC, normal controls; NES, Normalised Enrichment Score; KEGG, Kyoto Encyclopediaof Genes and Genomes; PPAR, peroxisome proliferator-activated receptor.
We then compared the expression levels of individual genes in each metabolic pathway in the CHB, LC, ACHD and ACLF groups with the NC group and observed robust changes in gene expression across ACLF development (figure 4B). An increase in the expression of metabolic genes was observed compared with other disease stages, most noticeably genes in the PPAR and mTOR signalling pathways, which further indicated that lipid metabolic disorders might play a key role in ACLF development. Additionally, ‘complement and coagulation cascades’ and ‘glycosphingolipid biosynthesis’ were significantly upregulated among metabolic pathways in patients with coagulation failure (online supplemental figure 4). Patients with cerebral dysfunction showed disorder in the ‘relaxin signalling pathway’, ‘inositol phosphate metabolism’ (online supplemental figure 5).
Immune-metabolism signatures associated with ACLF development
We performed four pairwise comparisons between disease stages (starting from NC, CHB, and LC to ACHD and ACLF) to identify the key molecules that were differentially expressed and related to ACLF pathophysiology. The frequencies of appearance of the top 50 DEGs in the four comparisons were counted (figure 5A). The top two DEGs (thrombospondin 1, THBS1; MER tyrosine kinase, MERTK) were identified, indicating that they were significantly differentially expressed in most comparisons among groups. Furthermore, semaphorin 6B (SEMA6B) displayed the maximum log2-fold change in expression, and PPAR gamma (PPARG) produced the minimum adjusted p value in the comparison of the ACLF and NC groups (figure 5B). The interaction network-based functional enrichment analysis showed that these four genes were involved in the innate immune response, adaptive immune response, complement activation, fatty acid oxidation and reactive oxygen species metabolism, suggesting that these molecules reflect dysregulated pathways that underlie the virus-based immune-metabolism disorder during the overall development of ACLF (figure 5C,D).

Immune-metabolism signatures associated with ACLF development. (A) The frequencies of the top 10 genes appearing in the top 50 DEGs of the four comparisons (ranked by adjusted p value). (B) The top 10 genes are shown in the volcano plot of the ACLF versus NC groups. Significantly differentially expressed genes (|log2-fold change|>1.5; adjusted p value <0.05) are shown in red or blue. The node size represents the frequency of key genes. (C) String network showing genes interacting with the top four genes (MER tyrosine kinase, MERTK; semaphorin 6B, SEMA6B; thrombospondin 1, THBS1 and PPAR gamma, PPARG). The colour represents the log2-fold change in the ACLF group compared with the NC group. (D) Significantly enriched pathways for the genes interacting with the top four genes (MERTK, SEMA6B, THBS1, PPARG, p<0.05). (E) Expression of four key genes in the sequencing group. Box plot, medians with (p25, p75). The false discovery rate was calculated using the Benjamini-Hochberg procedure. #No Significance, *adjusted p value <0.05, **adjusted p value <0.01, ***adjusted p value <0.001. The number of mRNA sequencing (n=20/10/10/10/15, ACLF/ACHD/LC/CHB/NC groups, respectively). (F) The qRT-PCR validation of four key genes (n=110/60/60/60/45, ACLF/ACHD/LC/CHB/NC groups, respectively). Plot, medians with (p25, p75). Mann-Whitney U test, #No Significance, *p<0.05, **p<0.01 and ***p<0.001. (G) Immunohistochemistry staining for four key proteins involved in ACLF development in patients from the HBV-ACLF, LC and NC groups. ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; CHB, chronic hepatitis B; DEGs, differentially expressed genes; HBV, hepatitis B virus; LC, liver cirrhosis; NC, normal controls.
The four key molecules related to immune-metabolism disorders associated with ACLF progression were validated in the external validation group. The clinical characteristics of the patients in the external validation group were similar to the patients in the sequencing group (online supplemental table 17). The qRT-PCR results showed increased expression of the four development-related genes during the progression from CHB/LC to ACHD and ACLF, consistent with the gene expression detected in the sequencing samples (figure 5E,F). IHC validation in liver tissues from NC and patients with LC and ACLF showed that the expression of the four proteins encoded by these disease development-related genes increased during disease progression from NC and LC to ACLF (figure 5G). These data confirmed the results of our transcriptomic analysis and suggested that the afore-mentioned genes should be further explored as potential diagnostic and prognostic biomarkers for ACLF.
Validation of four potential biomarkers in rat ACLF models
The four potential biomarkers were also validated in liver tissues from rats with ACLF using qRT-PCR and IHC to further confirm their specificity in patients with ACLF and potential research value. The rat ACLF model was induced by injecting D-galactosamine and lipopolysaccharides into porcine serum-treated rats (figure 6A, top panel). The analysis of liver function showed significantly increased levels of alanine aminotransferase, aspartate aminotransferase, total bile acid, TB and γ-glutamyl transpeptidase in ACLF rats (figure 6A, bottom left panel). H&E and Masson’s trichrome staining revealed the typical histology of cirrhosis-based liver failure, including extensive hepatocyte necrosis, an abnormal hepatic lobule structure, collagen accumulation, fibre hyperplasia and nodule formation (figure 6A, bottom right panel).

Validation of potential immune-metabolism biomarkers of ACLF development in a rat ACLF model. (A) ACLF was induced in porcine serum-treated rats through the injection of D-galactosamine and lipopolysaccharides. Top panel, schematic showing the design of the ACLF rat model. Bottom left panel, changes in six typical biochemical markers of liver function observed among the groups. Bottom right panel, pathological staining (H&E and Masson’s trichrome) of liver tissues (bar=250 µm). (B) The quantitative reverse transcription-PCR measurement of the expression of four key molecules in liver tissues (n=8/8/16, NC/LC/ACLF, respectively). (C) Images of IHC staining for four key molecules in rat liver tissues (bar=50 µm). Plot, means with SD. Kruskal-Wallis test, *p<0.05, **p<0.01, ***p<0.001 and ****p<0.001. ACHD, acute-on-chronic hepatic dysfunction; ACLF, acute-on-chronic liver failure; CHB, chronic hepatitis B; COSSH, Chinese Group on the Study of Severe Hepatitis B; DEGs, differentially expressed genes; HBV, hepatitis B virus; LC, liver cirrhosis; NC, normal controls.
Four potential biomarkers related to immune-metabolism disorders associated with ACLF development were validated by qRT-PCR in liver tissues from ACLF rats, and the results showed gradually increasing expression of MERTK, SEMA6B and THBS1 from NC and LC to ACLF rats, with the exception of PPARG (figure 6B). IHC staining showed gradually increasing expression of the MERTK, SEMA6B, THBS1 and PPARG proteins from NC and LC to ACLF rats (figure 6C). These results further confirmed the specificity of these markers in patients with HBV-ACLF.
Discussion
In pathophysiology, the interactions between virus exacerbation, immune processes and inflammatory processes create a complex network, many aspects of which remain unclear. Several retrospective studies and small cohort studies have reported evidence that collectively suggests that HBV exacerbation may induce excessive immune responses in patients with chronic hepatitis and/or LC.14 In the present study, the significantly increased expression of interferon-related, monocyte-related, neutrophil-related, inflammation-related and dendritic cell-related modules, and the significantly decreased expression of T cell-related, B cell-related and NK cell-related modules were observed in patients with ACHD, implying that the innate immune system was activated and that the adaptive immune system was exhausted during the evolving disease course starting from CHB or LC and developing into ACLF. Many viral processes participated in progression at all disease stages, indicating that excessive immune responses may be triggered by acute HBV exacerbation in patients with ACHD. Immune process dysregulation in patients with HBV-ACLF was more evident than inflammatory process dysregulation, in contrast to the main pathogenic pathway involving the systemic inflammatory response in patients with alcoholic liver disease-related ACLF.5
In this study, we also observed prominent metabolic alterations (including lipid metabolism, fatty acid metabolism, autophagy and oxygen homoeostasis) across all stages of ACLF development. A recent study further confirmed that metabolic dysregulation is associated with increased mortality and disease severity in patients and is related to extrahepatic organ failure,15 indicating that viral, immune and metabolic processes drive the development and progression of HBV-ACLF (figure 7).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proposed mechanism of HBV-ACLF development and progression. HBV-ACLF may be aggravated by an excessive immune response triggered by HBV exacerbation during the initial stage. Subsequent inflammatory responses and metabolic disorders result in multiorgan failure. The immune-metabolism disorder triggered by HBV exacerbation may be an important mechanism that aggravates HBV-ACLF. ACLF, acute-on-chronic liver failure; HBV, hepatitis B virus; IL, interleukin.
This study highlights the importance of immune-metabolism disorder in the pathogenesis of HBV-ACLF. A few studies have suggested that approaches designed to ameliorate immune dysfunction might be a viable strategy for the treatment of HBV-ACLF.16 Stem cell transplantation significantly increases the survival of patients with HBV-ACLF by exerting paracrine effects on the regulation of immune responses.17 As shown in our previous study, the paracrine effects of stem cells improve metabolism and promote regeneration through immune regulation in pigs.9 Taken together, treatments targeting immune regulation and metabolic balance represent a potential strategy for HBV-ACLF management, consistent with the pivotal role of immune-metabolism disorder observed in this study. However, an understanding of the precise roles of the immune-metabolism axis in ACLF development and progression requires additional proteomic and metabolomic evidence.
Furthermore, the identification of immune-metabolism biomarkers for ACLF may not only improve clinical practice but also provide molecular insights into the pathophysiological basis of this disease. Although we and other researchers have reported that serum levels of macrophage inflammatory protein 3α, K18 and cK18 can predict the severity of ACLF,18 19 the discovery of these biomarkers was based on common laboratory indicators or low-throughput techniques, which do not reflect the complicated molecular pathogenesis of the development, progression and outcomes of HBV-ACLF.6 In contrast, our study was based on the large prospective COSSH-ACLF cohort covering four evolving disease stages starting from CHB and progressing to LC, ACHD and ACLF. Our clinical manifestation-directed analysis of the functional synergy also enabled the discovery of four potential biomarkers (MERTK, PPARG, SEMA6B and THBS1) reflecting the virus-based immune-metabolism disorder in patients with ACLF. MERTK has been shown to suppress the innate immune response to microbes in patients with ACLF.20 PPARG has been reported to be significantly associated with steatosis and fibrosis, indicating that PPARG may be the target of cirrhosis and lipid metabolism.21 THBS1 has been documented to be a regulator of transforming growth factor beta (TGF-β) mediated suppression of liver regeneration.22 The validation with qRT-PCR and IHC in PBMCs and liver tissues from patients with HBV-ACLF and in liver tissues from ACLF rats further confirmed their important roles and indicated their potential to serve as biomarkers of HBV-ACLF pathogenesis. Our strategy of stratified random sampling and greater sequencing depth also reduced selection bias and ensured high-quality data. The detailed mechanisms should be clarified in future studies, particularly in a non-HBV ACLF population and ACLF animal models.23
In summary, HBV-ACLF has characteristic transcriptomic features in which an excessive immune response triggered by HBV exacerbation drives CHB and/or LC to progress into ACLF. Subsequent immune dysregulation, including innate immune activation and adaptive immune suppression, induces metabolic disruption. These findings highlight the importance of immune-metabolism disorder as a core axis of disease development and progression in HBV-ACLF, which may direct the development of new treatment strategies targeting HBV and immune-metabolism disorder to reduce the high-mortality rate of HBV-ACLF.
Data availability statement
Data, that is raw reads from RNA-seq experiments in fastq files, are available in a public, open access repository without restrictions on the use or distribution of the data. The Sequence Read Archive database project accession number for accessing the data produced for this study is PRJNA548207 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA548207/).
Ethics statements
Patient consent for publication
Ethics approval
The study protocol was approved by the Clinical Research Ethics Committee of the First Affiliated Hospital, Zhejiang University School of Medicine (No. 2011-13). All patients and volunteers were well informed, and written consent was obtained from the study subjects or the legal surrogates of the patients before enrolment.
Acknowledgments
The authors thank Dr. Vicente Arroyo (Liver Unit, Hospital Clinic, University of Barcelona, Spain) for providing helpful comments on preparing the manuscript.
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Twitter @lijun2009
Correction notice This article has been corrected since it published Online First. The provenance and peer review statement has been included.
Contributors JL (first author), XL, JJ and LY contributed equally. The study was designed by JL (last author) and XC. The manuscript was written by JL (last author), XC, JT, JL (first author), XL and DW. The experiments and data analysis were performed by JL (first author), XL, JJ, LY, JX, DS, YL, JL (8thauthor), KR, HH, JZ, PC, HY, JL (14th author), TW, LJ, PY, TL, HZ, SS, BG, XZ, QC, JC, XX, JH (26th author), SH, JH (28th author), SX, DW, JT, XC and JL (last author). JL (Last author) supported and supervised the project. All authors were involved in the critical revision of the manuscript.
Funding This study was supported by the National Natural Science Foundation of China (81830073), the National S&T Major Project of China (2017ZX10203201), the Zhejiang Provincial and State’s Key Project of Research and Development Plan of China (2016YFC1101303/4 and 2017C01026) and the National and Zhejiang Provincial special support programme for high-level personnel recruitment (Ten-thousand Talents Programme).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.